
[Back to Higher classification of Delphacidae]
[Back to A checklist of New World delphacid species]
Contents
Family Delphacidae Leach, 1815
Subfamily Plesiodelphacinae Asche, 1985
Tribe Plesiodelphacini Asche, 1985
Distribution: Neotropical (except 1 species Japan)
The Plesiodelphacini are specialized planthoppers with all known New World species on Heliconia (usually in the whorl of unopened leaves), and Burnilia japonica is on Alpinia intermedia (ginger family, ‘shell ginger’) .
Composition
Tribe Plesiodelphacini (subfamily Plesiodelphacinae is composed of a single tribe)
Burnilia Muir and Giffard, 1924 (Type species Delphax pictifrons Stål, 1864; 6 species Neotropical and Japan)
Burnilia belemensis Muir, 1926 -?Belize, Brazil, Venezuela
Burnilia heliconiae Muir, 1926 – Guiana
Burnilia japonica Asche, Hayashi & Fujinuma, 2016 – Japan (Kyushu)
Burnilia longicaput Muir, 1926 – Brazil, Guyana
Burnilia pictifrons (Stål, 1864) – Belize, Ecuador, Honduras, Mexico, Nicaragua
= Delphax pictifrons Stål, 1864
= Proterosydne pictifrons (Stål, 1864); new comb. by Crawford, 1914: 571.
= Burnilia pictifrons (Stål, 1864); new comb. by Muir and Giffard, 1924: 7.
Burnilia spinifera Fennah, 1945 – Dominica, Guyana, St. Vincent, Trinidad
= Burnilia spinifera spinifera Fennah, 1945; subspecies by implication Fennah, 1959: 251.
Burnilia spinifera antillana Fennah, 1959 – Dominica, St. Vincent
Burnilia williamsi Muir, 1926 – Brazil



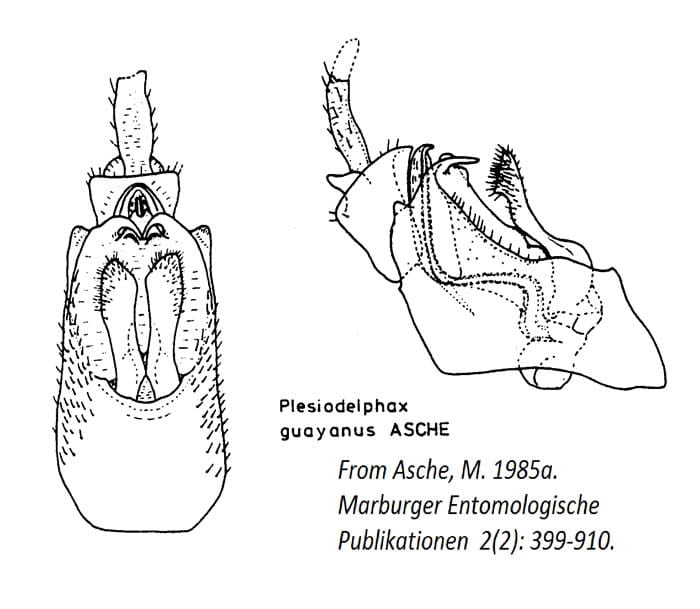
Plesiodelphax Asche, 1985 (Type species Plesiodelphax guayanus Asche, 1985 by monotypy; northeastern South America)
Plesiodelphax guayanus Asche, 1985b – Brazil, French Guiana
(See Line drawings below, I don’t have photos of Plesiodelphax)
Concise features of the tribe: (compiled from Asche 1985b, 1990; compare with features of the Asiracinae here) (I’ll update with Asche 1985a later).
1. Tymbal sexually dimorphic (Apomorphic versus Asiracinae).
-. Abdominal sternite 2 with elongate apodemes (part of tymbal) (Apomorphic versus Asiracinae, apodemes secondarily reduced in advanced lineages).
-. Central plate present in male drumming organ.
2. Calcar somewhat flattened (not tectiform), circular in cross-section, with large teeth along trailing margin (apomorphic versus Vizcayinae and Asiracinae).
3. Apical spines of hind tibiae differentiated into 2 groups (apomorphic versus Vizcayinae and Asiracinae).
-. Apical spines of basitarus of hind leg not in uniform row (apomorphic versus Vizcayinae and Asiracinae).
4. Ring-like rupture at posterior pole of egg chorion.
5. Aedeagus with sclerotized sperm-conducting tube (plesiomorphic).
-. Flagellum of aedeagus reduced or absent (absent is apomorphic)
6. Anal tube and thecal base in close contact (suspensorium absent) (plesiomorphic)
7. “Symbiont x” presumed present (plesiomorphic)
8. Hind wings with M and Cu fused over most of their length
9. Hind wings with 4 veins from nodal line (i.e., M unbranched or branched after nodal line) (shared with higher delphacids)
10. Two lateral teeth on hind tibiae (apomorphic versus Asiracinae, shared with most higher Delphacidae)
11. Genal carina extending from lateral frontoclypeal edge to anterior edge of antennae (shared with all higher delphacids).
12. Median carina of vertex obsolete (or nearly so) (synapomorphy)
13. Antennal segments 1 & 2 of differing length (shared with higher delphacids)
-. Antennal sensory structures in more-or-less regular rows (reduced to 16, although I have not counted; shared with all higher delphacids).
Asche (1985a, 1990) considers this subfamily to be relatively derived , whereas Urban et al. (2010) placed the subfamily basad of the Kelisiini
Asche et al. (2016: 75-76) described the systematic placement of the subfamily as follows
“Due to the display of a cross-sectional circular post-tibial spur with well separated conical teeth Muir and Giffard (1924) placed Burnilia into the tribe Alohini Muir, 1915, still listed here in Metcalf (1943). Asche (1985a) described a second genus close to Burnilia, viz., Plesiodelphax with the type species Plesiodelphax guayanus Asche, 1985 from Brazil (type locality: Porto 14 de Mayo) and French Guiana. Moreover, Asche (1985a) removed Burnilia from the Alohini (the latter being integrated as junior synonym into the Delphacini: Asche 1985b), and established the subfamily Plesiodelphacinae for Burnilia and Plesiodelphax, based on characters regarded as autapomorphies (concerning hind wing venation, the special carination of the vertex, the arrangements of distal spine at the first post-metatarsus, and in the special configuration of the male genitalia, as described and discussed in Asche 1985b). The display of the post-tibial spur was rather regarded as “kelisioid” than as “alohinid” (Asche 1985a). In his phylogeny of Delphacidae, Asche (1985b), placed the Plesiodelphacinae above the level of Stenocraninae as the sistergroup of Delphacinae (see also Asche 1990). Mainly based on larval morphology, Emeljanov (1995) distinguished only three subfamilies of Delphacidae: Asiracinae, Ugyopinae and Delphacinae. The latter comprises seven tribes including Plesiodelphacini which were placed in his cladogram above the level of Stenocranini and considered as the sistergroup of a clade Tropidocephalini – Saccharosydnini plus Delphacini, – congruent with the hypothesis of Asche (1985b). However, Emeljanov ́s tribal classification is not fully adopted here, with consequence that plesiodelphacine taxa are still regarded as separate subfamily as suggested by Asche (1985b, 1990). The higher classification of Delphacidae has also been addressed by Hamilton (2006) who interpreted and supplemented mainly Emeljanov’s data (Emeljanov 1995). Hamilton (2006) moved most subfamilies and tribes with the exception of Plesiodelphacinae to a single subfamily “Delphacinae”. For Plesiodelphacinae he referred to Bartlett (2005) who reported about possible relationships of this group to Asiracinae. Urban et al. (2010) investigated the phylogeny of Delphacidae on the basis of molecular data and implications revealed from host plant associations, providing the first substantive molecular phylogeny of this family. According to their data, Plesiodelphacinae would possibly have derived from a level basally of Kelisiinae and Stenocraninae; however, as this hypothesis would have a substantial impact on the interpretation of morphological data as suggested earlier by Asche (1985b) and Emeljanov (1995)…”
Recognition





Selected references
Asche, M. 1985a. A new subfamily, genus and species of Delphacidae from South America: Plesiodelphacinae subfam. nov., Plesiodelphax guayanus gen. et spec. nov. (Homoptera Fulgoroidea). Marburger Entomologische Publikationen 1(10) : 219-240.
Asche, M. 1985b. Zur Phylogenie der Delphacidae Leach, 1815 (Homoptera: Cicadina: Fulgoromorpha). Marburger Entomologische Publikationen Marburger Entomologische Publikationen 2(1):1-398 AND 2(2): 399-910.
Asche, M. 1990. Vizcayinae, a new subfamily of Delphacidae with revision of Vizcaya Muir (Homoptera: Fulgoroidea) – a significant phylogenetic link. Bishop Museum Occasional Papers 30: 154-187.
Asche, M., M. Hayashi and S. Fujinuma. 2016. Enigmatic distribution: first record of a hitherto New World planthopper taxon from Japan (Hemiptera, Fulgoroidea, Delphacidae, Plesiodelphacinae). Deutsche entomologische Zeitschrift 63(1): 75-88.
Bartlett. C. R. 2005. Progress in the phylogeny of the Delphacidae using molecular and morphological tools. In: Abstracts of talks and posters, 12th International Auchenorrhyncha Congress, University of California, Berkeley, 7–12 August 2005, S[ymposia]: 29–30.
Bartlett, C. R., L. B. O’Brien and S. W. Wilson. 2014. A review of the planthoppers (Hemiptera: Fulgoroidea) of the United States. Memoirs of the American Entomological Society 50: 1-287.
Crawford, D. L. 1914a. A contribution toward a monograph of the homopterous insects of the family Delphacidae of North and South America. Proceedings of the United States National Museum 46: 557-640, plus 6 plates.
Emeljanov, A. F. 1995. On the question of the classification and phylogeny of the Delphacidae (Homoptera, Cicadina). Entomologicheskoye Obozrenie 74(4): 780–794. [In Russian] [Translated into English in Entomological Review 1996 75(9): 134–150.]
Fennah, R. G. 1945. The Fulgoroidea, or lanternflies, of Trinidad and adjacent parts of South America. Proceedings of the United States National Museum, Washington, 95: 411–520, plates 7–17. doi: 10.5479/si.00963801.95-3184.411
Fennah, R. G. 1959. Delphacidae from the Lesser Antilles (Homoptera: Fulgoroidea). Bulletin of the British Museum (Natural History) Entomology 8(6): 245–265.
Hamilton, K.G.A. 2006. The planthopper genus Stenocranus in Canada: implications for classification of Delphacidae (Hemiptera). Canadian Entomologist 138: 493–503. doi: 10.4039/n06-805.
Kirkaldy, G. W. 1907i. Leafhoppers. Hemiptera Homoptera. Report of work of the Experiment Station of the Hawaiian Sugar Planters’ Association. Division of Entomology bulletin. 4: 60-66.
Leach, W. E. 1815a. Entomology. The Edinburg encyclopedia; conducted by David Brewster 9: 57-172. (family Delphacidae page 125).
Metcalf, Z. P. 1943. General Catalogue of the Hemiptera. Fascicle IV, Fulgoroidea, Part 3, Araeopidae (Delphacidae). Smith College, Northhampton, Massachusetts.
Muir, F.A.G. 1915b-e. A contribution towards the taxonomy of the Delphacidae. Canadian Entomologist 47: 208-212, 261-270, 296-302, 317-320.
Muir, F.A.G. 1926b. Contributions to our knowledge of South American Fulgoroidea (Homoptera). Part I. The Family Delphacidae. Experiment Station of the Hawaiian Sugar Planters’ Association, Entomological Series, Bulletin 18: 1-51, plates 1-5.
Muir, F.A.G. and W. M. Giffard. 1924a. Studies in North American Delphacidae. Bulletin of the Hawaiian Sugar Planters Association Division of Entomology 15: 1-53.
Stål, C. 1864. Hemiptera mexicana enumeravit speciesque novas descripsit. (Continuatio). Stettiner Entomologische Zeitung 25: 49–86.
Urban J. M., C. R. Bartlett and J. R. Cryan. 2010. Evolution of Delphacidae (Hemiptera: Fulgoroidea): combined-evidence phylogenetics reveals importance of grass host shifts. Systematic Entomology 35(4): 678-691.
